Hardy-Weinberg Equilibrium

G. H. Hardy
By Unknown - A mathematician's apology, Public Domain, https://commons.wikimedia.org/w/index.php?curid=3778423
Today’s post is not about useful data, but useful theory. This is not my field, but it is fun. The Hardy-Weinberg Equation is a remarkably simple bit of theory that comes from the logical relationships between genes and genotypes. Given some assumptions, you can readily calculate either the gene frequencies if the genotypes are known, or the genotype distribution if the gene proportions are known. Strictly speaking, the relationship is between the gene frequencies of the parents, and the genotypes of their offspring, but when Hardy-Weinberg equilibrium is in effect, the frequencies of both are stable with time.
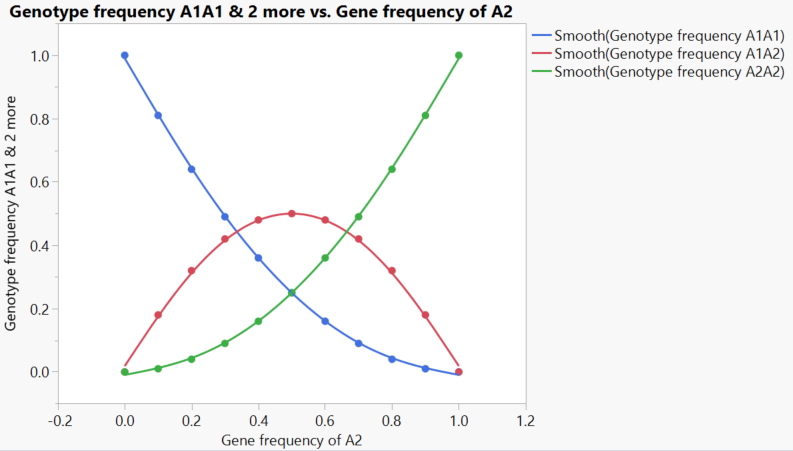
The simplest case of two genes
The assumptions are important. You won’t get the expected gene frequencies if they are violated, but the deviations usually tell you something interesting.
- Large population
- Random mating
- Gene frequencies are equal in the sexes
- There is no migration, gene flow, admixture, mutation, or selection
- Plus some technical details about how gametogenesis occurs
There some useful things that come directly from the structure of this relationship. For example, in the case of deleterious recessive mutations, heterozygotes [carriers] will be vastly more common than homozygotes, who actually express whatever the mutation causes.

If A1 is a deleterious recessive, the number of heterozygotes can be many orders of magnitude greater than those with two copies who suffer whatever the condition is
This particular case is pretty sensitive to the effects of non-random mating, which happens pretty frequently. However, one of the most fascinating things about the Hardy-Weinberg relationship is that it works gene by gene. Selection can be occurring in other genes, but for any that are not, the Hardy-Weinberg equilibrium will hold as long as the other assumptions are met.
The example is a simple one of two genes in a diploid organism, but the relationship can be readily generalized to any number of alleles at a locus or polyploidy.
In somes cases where the Hardy-Weinberg equilibrium conditions are not met, this changes from a static relationship to a dynamic one, with predictable changes in gene frequencies occurring over time. For example, if the males and females have different gene frequencies to start, their offspring will oscillate around equilibrium for a few generations until settling down to the the equilibrium point.
Another interesting result occurs when mating is not random. For example, if like phenotypes are more likely to mate [a not uncommon thing!] then the proportion of homozygotes goes up. If you happen to be a carrier of a deleterious recessive, which most of us are, this is a bad thing, as it increases the chance of the offspring inheriting two copies of the deleterious recessive. This is why it is a bad idea to have kids with your cousin.
Even for so basic an overview, there is a lot of useful information you get out of something so simple, and this is foundational for understanding other things in population genetics.


Comments ()